Dr. Giovanni Chetta tarafından düzenlendi
hücre dışı ağ
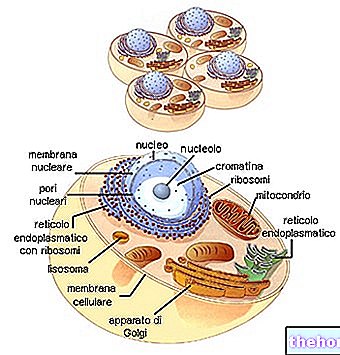
Bu nedenle ECM, proteinlerin, PGS'lerin ve GAG'ların, yapısal destek ve her doku ve organik aktivitenin düzenlenmesi dahil olmak üzere sayısız işlev sağladığı çok karmaşık bir ağ olarak düşünülebilir. Küresel hücresel homeostaz, ECM'de hücre içinde veya dışında ortaya çıkabilen ve gelişebilen bir mekanizmalar kompleksi olarak düşünülmelidir; ikinci durumda, hücre ara veya nihai hedefi temsil edebilir. Hücre dışı bileşenler, hücresel yapı iskelesi için fiziksel destek yapılarını temsil etmenin yanı sıra, hem hücre içi çevre hem de organlar ve sistemler ile ilgili hayati süreçlerin başlatılması, geliştirilmesi ve sonlandırılması için gerçek alanlar olarak işlev görür. Milyonlarca ve milyonlarca bilgiyi uzaktan bile üretebilen, modüle edebilen, değiştirebilen ve yayabilen sonsuz bir biyokimyasal ağla karşı karşıyayız.
Vücudun her hücresi, dokuların statik ve dinamik mimarisi üzerinde "dramatik" etkilerle hem mekanik hem de kimyasal ve enerjik yönler altında ECM ile sürekli etkileşime girer. Örneğin fibroblastlar, belirli organik fonksiyonel bölgenin gerektirdiği biçimlerde ve miktarlarda sıkıştırabilmek ve hazırlayabilmek için sürekli olarak işleyerek ürettikleri kollajen üzerinde büyük ölçüde etki eder. İki küçük embriyonik doku parçası birbirinden uzak fakat aynı kolajen jel kültürüne yerleştirilirse, önce iki kütüğü birbirine bağlayan mükemmel şekilde hizalanmış neokollajen liflerinin oluşumuna tanık oluruz. Daha sonra, fibroblastlar, birikimlerini kontrol ederek ve sırayla izlenerek, neo-kollajen lifler boyunca iki parçadan dışarı göç eder. Bu fonksiyonel sinsityum muhtemelen ECM'nin rejeneratif süreçleri sırasında mevcuttur ve çeşitli hayati doku fonksiyonlarının gerektirdiği sabit varyasyonlarla fizyolojik koşullar altında başa çıkarak kendi kendini düzenleyebilen çok yıllık bir fonksiyonel süreklilik oluşturur (Albergati, 2004).
P. A. Bacci'ye göre, interstisyel matris, her şeyden önce madde ve enerji arasındaki alışverişin gerçekleştiği yer olan hayati reaksiyonların anasını gerçekten temsil eder. Tüm dokular kapalı değil açık sistemlerde birbirine bağlı ve işlevsel olarak entegredir; aralarında biyokimyasal, biyofiziksel ve elektromanyetik mesajlardan yararlanarak, yani çeşitli enerji biçimlerini kullanarak hem yerel hem de sistemik olarak gerçekleşebilen sürekli alışverişler gerçekleşir. Hücre dışı interstisyel boşluğun iyonik bileşimi, yalnızca değiş tokuşlara ve yaşama izin veren değil, aynı zamanda her hücrenin gen ekspresyonu üzerinde de etki eden temel bir madde oluşturur.
F. G. Albergati'nin belirttiği gibi, hücre ve hücre dışı matris, yalnızca görünüşte ayrı olan ve doğru ve sinerjik bir şekilde çalışmak için her an zorunlu olarak tüm yaşam süresi boyunca etkileşime girmesi gereken iki dünyayı temsil eder. Bu, olağanüstü bir dizi sinyalin ardından eşit derecede inanılmaz bir dizi moleküler-biyolojik aktivite gerektirir.
MEC'in yeniden yapılandırılması
ECM, hem fizyolojik hem de patolojik koşullarda, hem kendi içinden ("metalloproteazların eylemi yoluyla) hem de hücrelerden gelen işlevsel taleplere dayalı olarak çok yıllık ve sürekli morfo-fonksiyonel "yeniden şekillenme" içinde bir yapı olarak düşünülmelidir. "yapışmanın sayısız protein fraksiyonunun eylemi için). Azaltılmış veya eksik ECM yeniden şekillenme kapasitesi hücre için öldürücüdür. Gördüğümüz gibi, tüm hücresel işlevler, ECM'nin yapılarının karşılık gelen işlevi sayesinde ifade edilir ve patolojik süreçler, ECM'nin modifikasyonlarına birincil veya ikincil olabilir.
ECM yeniden şekillenmesinin açık örnekleri, dermal-hipodermik katmanların onarımıdır (bu işlem, kesin matris bozunma dizileri, bu bölgedeki belirli hücrelerin göçü, fibronektin, fibrinler ve büyük miktarlarda tip III kollajenden oluşan geçici bir matrisin sentezini gerektirir, bileşenlerinin fonksiyonel restorasyonu ve sonuç olarak dokunun yapısal restorasyonu için temel olan geçici matrisin yeniden şekillenmesi aşaması) ve neoanjiyogenez (onkolojik dahil olmak üzere çeşitli patolojik durumlarda dokularda ve organlarda yeni kılcal damarların oluşumunun fizyolojik süreci) . Neoanjiyogenez, örneğin kalp kası veya periferik dolaşım gibi iskemik dokularda doğru kan akışını yeniden sağlamak ve ayrıca, örneğin tümör alanında, süreci inhibe etmek amacıyla terapötik amaçlar için incelenir (Shishido ve diğerleri, 2003). )
NS metaloproteaz (MMP'ler), ECM'nin tüm proteinlerini ve proteoglikan bileşenlerini parçalama potansiyeli gösteren bir çinko ve kalsiyum içeren endopeptidaz ailesidir. İnterstisyel kollajene benzer dizilere sahiptirler ve hücre zarının dış tarafına inaktif (gerektiğinde aktive edilmiş) bir biçimde yerleştirilirler. Aktiviteleri, TIMP (MetalloProteazların Doku İnhibitörleri) adı verilen özel bir ajan tarafından inhibe edilir.
Geçmişte, doğal veya denatüre kolajeni (jelatin) denatüre etme özellikleri nedeniyle kolajenaz veya jelatinaz olarak anılırdı - Birkedal-Hansen. Bu enzimler, anjiyogenez, embriyogenez, enflamatuar tepkiler, ateroskleroz ve çok sayıda artritik hastalık (romatoid artrit dahil - Dieppe, 1995) dahil olmak üzere çok sayıda fizyolojik ve patolojik süreçte yer alır.
"Hücresel dengede hücre dışı matrisin önemi" ile ilgili diğer makaleler
- Fibronektin, Glukozaminoglikanlar ve Proteoglikanlar
- Hücre dışı matris
- Kollajen ve elastin, hücre dışı matristeki kolajen lifleri
- Hücre dışı matris ve patolojilerin değişiklikleri
- Bağ dokusu ve hücre dışı matris
- Derin fasya - Bağ dokusu
- Fasyal mekanoreseptörler ve miyofibroblastlar
- Derin fasya biyomekaniği
- Duruş ve dinamik denge
- Gerginlik ve sarmal hareketler
- Alt ekstremiteler ve vücut hareketi
- Makat desteği ve stomatognatik aparat
- Klinik vakalar, duruş değişiklikleri
- Klinik vakalar, duruş
- Postural değerlendirme - Klinik vaka
- Kaynakça - Hücre dışı matristen duruşa. Bağlayıcı sistem bizim gerçek Deus eski makinemiz mi?