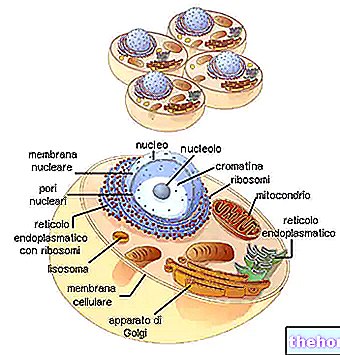
ORTAKLIK (VEYA "BİRİŞTİRME" VEYA "BAĞLANTI")
Şimdiye kadar Mendel çift hibrit (veya polihibrit) çaprazlamasını farklı alelik çiftlerin aslında farklı homolog kromozom çiftlerinde bulunduğunu varsayarak ele aldık.Fakat kromozom çiftlerinin sayısı türden türe farklılık gösterse de dar sınırlar içinde değişir (birkaç tür yaklaşık yüz kromozoma ulaşır), genlerin sayısı ise on binlerce olarak sayılabilir.
Mendel'in deneyleri için seçtiği karakterlerin hepsini bağımsız olarak ayırması (dolayısıyla polihibrit çaprazın F2'sindeki fenotiplerin dağılımındaki hesaplamaları karıştırmadan) şanslı bir şanstı.Eğer bitişik lokuslarda iki çift alel bulunursa, takip edecekse, dernek yasası olarak adlandırılacaktır.
Pek çok karakterin tek bir kromozomda lokusları olduğu ve bunların mayoz bölünmede bağımsız olarak ayrılan kromozom çiftleri olduğu bilindiğinde, ebeveynde bir kromozom üzerinde ilişkilendirilmişlerse iki karakter çiftinin ne sıklıkla meydana geldiği iyi anlaşılabilir. organizma, gamette ve dolayısıyla kendi genetik materyalini getireceği organizmada da eşit olarak ilişkili kalır.
Böylece, Mendel'in üçüncü yasasında ifade edilen bağımsızlığın "bir istisnasını" temsil ettiğini görüyoruz.
DEĞİŞİM VEYA "ÇAPRAZ GEÇİŞ" VE REKOMBİNASYON
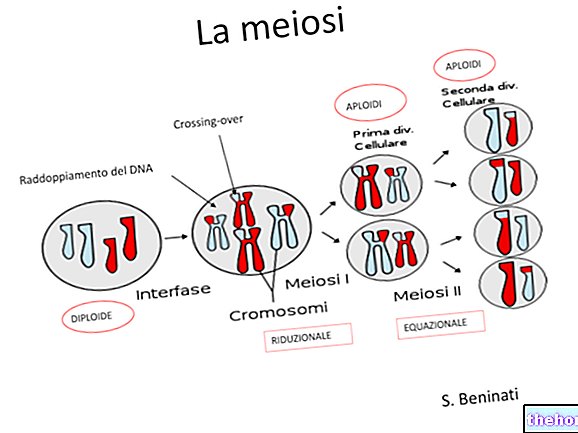
Mayozdan bahsetmişken, genetik materyalin karışmasının iki farklı anı olduğunu belirtmiştik: biri gametlerde kromozomların ayrılması ve bu Mendel tarafından gözlemlenendir.
Aslında önce gelen diğer an, her bir homolog kromozom çiftinin dört kromatidinin karşılıklı olarak aynı özellikleri değiştirdiği andır.Bu alışverişi takiben, aynı kromozom üzerinde ilişkili iki faktör gametlerde bağımsız olacaktır.Olasılık Bir değiş tokuşun gerçekleşmesi, bir ilk yaklaşımla, kromozomun uzunluğuyla orantılıdır ve daha uzun kromozomlarda bir değiş tokuştan daha fazlası olabilir.
Bu fenomen, mikroskop altında yeterli sayıda mayoz gözlenerek sitolojik olarak tespit edilebilir.
Yeniden eşleşme oranı, ebeveyn neslinde ilişkilendirilmiş herhangi iki karakterin F2'de farklı şekilde yeniden birleşme hızıdır.
Eğer iki lokus kesinlikle bitişik ise, bir kiazmanın onları ayırma olasılığı pratikte sıfır olacaktır. Rekombinasyon oranı şöyle olacaktır: n ° rekombinant. İki lokus iki farklı kromozom üzerindeyse, rekombinasyon oranı 0,5 olacaktır (P nesline katılan iki karakterin kendilerini F2'de rastgele bir arada bulması için eşit olasılık). Bu nedenle rekombinasyon oranı 0.0 ile 0.5 arasında değişebilir. Kromozom üzerindeki küçük mesafeler için mesafe ve rekombinasyon oranı doğru orantılıdır. Daha uzun mesafeler için, iki lokus arasında iki alışverişin gerçekleşmesi olasılığı vardır. Şimdi, iki değiş tokuşla ayrılan iki faktörün bir kez daha ilişkilendirildiği açıkça görülecektir. Bu noktada, lokusların uzaklığı ile rekombinasyon olasılığı arasındaki orantılılığın kaybolduğu açıktır.
Aynı kromozom üzerinde ilişkili bulunan lokuslar 'birleşim grupları' oluşturur. Çok uzak lokuslar, bağımsız olarak davranacak kadar değiş tokuşla ayrılma olasılığına sahip olabilir, ancak her biri daha düşük bir rekombinasyon oranıyla ara lokuslarla ilişkilendirilecektir.
Bir ilişki grubu içindeki birçok gen çifti arasındaki rekombinasyon oranları bilindiğinde, 'genetik haritaların' yapımı başlayabilir. İki gen (a ve b) arasındaki mesafenin rekombinasyon oranı ile ifade edildiğini ve a'nın üçüncü bir gen c'ye olan mesafesinin, b'ye olan uzaklığına göre toplam veya fark olabileceği akılda tutularak, bu mümkündür. bu birliktelik grubu, yani o kromozom içindeki genetik harita olacak karşılıklı mesafelerin bir haritasını yeniden oluşturmak.
Şimdi, genotipik karakterlerin fenotipik tezahürünü sınırlayan bazı kavramları genel olarak ele almalıyız.
Her şeyden önce, nüfuz etme ve ifade etme kavramları hakkında konuşacağız ve daha sonra gen eyleminin düzenlenmesi fenomenine özellikle dikkat edeceğiz.
PENETRANSI
Bir genin nüfuzu, kendisini fenotipte gösterme yeteneğini temsil eder. Penetrasyon, onu içeren 100 genotipten o karakteri gösteren fenotiplerin sıklığı sayılarak istatistiksel olarak ölçülür. 0.7 penetranslı bir özellik, genotipik frekansının %70'inde fenotipik olarak ortaya çıkan bir özelliktir.
İFADE
Dışavurumculuk, fenotipik tezahürün derecesinin nicel bir değerlendirmesidir.
GEN EYLEMİNİN DÜZENLENMESİ
Hücreler tüm enzimlerini ve proteinlerini aynı hızda ve aynı zamanda üretirler. Örneğin Escherichia coli hücreleri, beta-galaktosidaz enzimi sayesinde glikoz ve galaktoza parçalayabildikleri için laktoz disakkaritten enerji ve karbon atomları ile sağlanabilir. o hücrenin proteinlerinin %3'üne eşit olan yaklaşık 3.000 beta-galaktosidaz molekülü vardır; laktoz yokluğunda bakteri hücresi başına sadece bir beta-galaktosidaz molekülü olacaktır. Galaktosidaz, kullanılabildiği zaman yeni mRNA moleküllerinden sentezlenecektir. Enzim açısından zengin mutant E. coli suşları, laktoz olmadığında bile bilinmektedir: bu mutantlar, substratsız kalacak olan enzimi üretmek için gereksiz bir enerji ve malzeme tüketimine zorlandıklarından, normal hücrelere kıyasla dezavantajlıdırlar. Laktozda olduğu gibi enzim miktarında artışa neden olan maddelere indükleyici, enzimlere indüklenebilir denilecektir. Diğer maddeler, bunlar da belirli bir şekilde belirli enzimlerin üretimini indükler. Ayrıca, örneğin, karbon ve amonyum (NH3) içeren tüm amino asitlerini oluşturabilen E. coli'de, belirli bir amino asidin (örneğin histidin) kültür ortamında bulunması, ilgili tüm enzimlerin üretimini bloke eder. amino asidin kendisinin biyosentezi: bu enzimlerin bastırılabilir oldukları söylenecektir.Bakteriyel hücrelerde mRNA molekülleri, oluşumlarından kısa bir süre sonra yıkılırlar ve bu nedenle mRNA üretimini kontrol etmek aynı zamanda enzimatik sentezi kontrol etmek anlamına gelir. zaman.
OPERON
Bakteri hücresinin kendi enzim üretimini nasıl kontrol edebildiğini açıklamak için Jacob ve Monod operon hipotezini formüle ettiler; operon, işlevsel olarak ilişkili ve bir DNA dizisi boyunca süreksizlik olmaksızın hizalanmış birkaç gen tarafından oluşturulur.Operon üç farklı gen tipinden oluşur: mRNA'nın oluşumunun başladığı promotör; kontrolün uygulandığı operatör, enzimleri veya diğer proteinleri kodlayan bir veya daha fazla yapısal gen Beta-galaktosidaz sisteminde, operon, beta-galaktosidaz için olana ek olarak, aynı zamanda yapısal kodlayan iki başka gen içerir. laktoz metabolizmasında yer alan diğer enzimler. Bu genler birbirine bitişiktir ve aynı DNA sarmalı boyunca birbiri ardına tek bir mRNA molekülüne kopyalanırlar.Bu şekilde üretilen mRNA molekülleri çok kısa bir süre aktif kalır, ardından spesifik enzimler tarafından yok edilirler.
Operonun aktivitesi sırayla başka bir gen tarafından kontrol edilir, regülatör, bu da operondan uzak olabilir: bu regülatör, operatör genindeki DNA'ya bağlandığı görünen, represör adı verilen bir proteini kodlar. promotör ve yapısal genler arasındaki ilişki aslında mRNA üretimini bloke eder.
Bastırıcı sırayla kontrol edilir ve kontrol bir "sinyal" maddesi vasıtasıyla gerçekleştirilir. İndüklenebilir enzimler söz konusu olduğunda bu madde "indükleyicidir. İndükleyici", şeklini değiştirerek baskılayıcı moleküle bağlanır, böylece artık DNA'ya uyum sağlayamaz: bu durumda, promotör ile yapısal genler arasında baskılayıcı olmadığı için baskılayıcı, mRNA moleküllerini ve bunlardan protein moleküllerini oluşturabilir. İndükleyici kaynağının tekrar tükenmesiyle, regülatör kontrolü yeniden kazanacak ve bu da yeni mRNA'nın, dolayısıyla yeni proteinlerin üretimini durduracaktır.Beta-galaktosidaz sisteminde indükleyici, laktoz veya buna çok benzer bir maddedir.Türev: onlar enzimlerin biyosentezine izin vermek için onu inaktive ederek baskılayıcıya katılır. Bastırılabilir enzimler durumunda, "sinyal" görevi gören madde bir koruyucu olarak hareket eder: baskılayıcı, yalnızca koruyucu ile birleştiğinde aktiftir. Bir düzine farklı enzimi içeren histidin sisteminde, tRNA'sı ile birleşen bu amino asit, korpresör histidindir.
ALLOSTERİK ETKİLEŞİMLER
Bir enzimin şeklini değiştirerek inaktivasyonunu içeren allosterik etkileşimler, bir hücrenin metabolik aktivitesini düzenlemenin farklı bir yolunu sağlar. Allosterik etkileşimler, operonun indüktör-baskılayıcı sisteminden daha doğru kontrole izin verir, ancak belirli bir maddenin biyosentezini ilk aşamadan - bir mRNA'nın üretiminden - hariç tutmak gibi yararlı bir sonuca ulaşmaz.
EUCARIUS'TA KONTROL SİSTEMLERİ
Bitkiler ve hayvanlar arasında operona benzer bir düzenleme sisteminin işlediğine ve baskın olduğuna inanmaya yol açan bazı gerçekler vardır.Bu organizmaların kromozomları, E.coli ve diğer prokaryotların kromozomlarından derinden farklıdır.Bunlardaki genlerin kontrolü. hücreler çok farklıdır.Mitozun mekanizması, belirli bir bitki veya hayvanın her hücresinin tüm bilgilere sahip olduğu şekildedir.
Döllenmiş yumurtada bulunan genetik. Bu nedenle, herhangi bir özel hücredeki genlerin çoğu, hücrenin ömrü boyunca etkisiz kalacaktır. Bu hücrelerdeki DNA her zaman proteinlerle ilişkilidir. Dolayısıyla, ökaryotlarda gen baskısının tam olarak bu ilişkiyi gerektirmesi mümkündür. DNA ve proteinler arasında.














.jpg)




.jpg)


