Polinükleotidin bilgileri ile polipeptidin bilgileri arasında bir yazışma olması için bir kod vardır: genetik kod.
Genetik kodun genel özellikleri şu şekilde sıralanabilir:
Genetik kod üçüzlerden oluşur ve dahili noktalama işaretlerinden yoksundur (Crick & Brenner,).
"Açık hücre çeviri sistemleri" kullanılarak deşifre edildi (Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Son derece dejeneredir (eşanlamlılar).
Kod tablosunun organizasyonu tesadüfi değildir.
Üçüzler "saçmalık".
Genetik kod "standart", ancak "evrensel" değil.
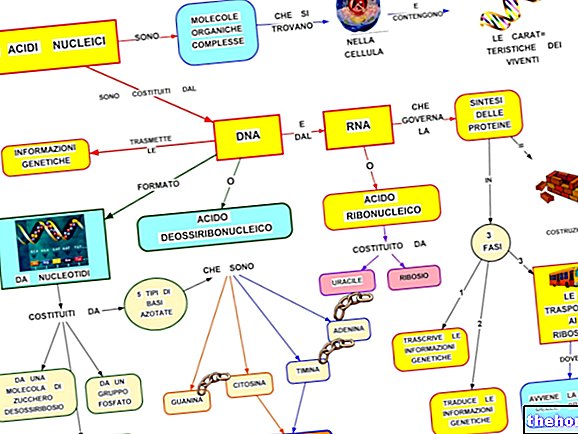
Genetik kod tablosuna bakıldığında, "RNAm'ın, ilgili nükleotid bazlarının A, U, G, C olduğu polipeptide çevirisini ifade ettiği unutulmamalıdır. Bir polipeptit zincirinin biyosentezi, translasyonudur. amino asit dizisindeki nükleotid dizisi.

RNAm'ın kodon adı verilen her bir baz üçlüsü, birinci baz sol sütunda, ikinci baz üst satırda, üçüncü baz sağ sütunda yer alır.Örneğin triptofan (yani Try) alalım ve buna karşılık gelen kodonun olacağını görüyoruz. sırayla, UGG olun. Aslında, ilk taban U, en üstteki tüm kutu sırasını içerir; bunda G, en sağdaki kutuyu ve kutunun kendisinin, yazılı Try'i bulduğumuz dördüncü satırını tanımlar. Benzer şekilde, tetrapeptid Lösin-Alanin-Arginin-Serína'yı (semboller Leu-Ala-Arg-Ser) sentezlemek için kodda UUA-AUC-AGA-UCA kodonlarını bulabiliriz.

Ancak bu noktada, tetrapeptidimizin tüm amino asitlerinin (triptofanın aksine) birden fazla kodon tarafından kodlandığına dikkat edilmelidir. Az önce bildirilen örnekte belirtilen kodonları seçmemiz tesadüf değildir Aynı tripeptidi CUC-GCC-CGG-UCC gibi farklı bir RNAm dizisi ile kodlamış olabilirdik.
Başlangıçta, tek bir amino asidin bir üçlüden fazlasına karşılık geldiği gerçeğine, eşanlamlılık fenomenini tanımlamak için kullanılan kodun dejenerasyon teriminin seçiminde de ifade edilen bir rastgelelik anlamı verildi. Öte yandan, bazı veriler, genetik bilginin farklı stabilitesine atıfta bulunulan eşanlamlıların mevcudiyetinin hiç de tesadüfi olmadığını göstermektedir.Bu, A + T / G + C oranının farklı bir değerinin bulunmasıyla da doğrulanmış gibi görünmektedir. evrimin farklı aşamalarında. Örneğin, değişkenlik ihtiyacının Mendelizm ve neo-Mendelizm kurallarıyla karşılanmadığı prokaryotlarda, A + T / G + C oranı artma eğilimindedir. gen mutasyonundan rastgele değişkenlik fırsatları.
Ökaryotlarda, özellikle tek bir organizmanın hücrelerinin hepsinin aynı kalıtsal mirası taşımasının gerekli olduğu çok hücreli hücrelerde, DNA'daki A + T / G + C oranı azalma eğilimi gösterir ve somatik gen mutasyonları olasılığını azaltır. .
Genetik kodda eşanlamlı kodonların mevcudiyeti, daha önce bahsedilen, RNAt'taki antikodonların çokluğu ya da olmaması sorununu gündeme getirmektedir.
Her amino asit için en az bir RNAt olduğu kesindir, ancak tek bir RNAt'ın tek bir kodona bağlanıp bağlanamayacağı veya eşanlamlıları fark etmeden tanıyabileceği (özellikle bunlar yalnızca üçüncü baz için farklılık gösterdiğinde) eşit derecede kesin değildir.
Antikodonların en az bir ve üçten fazla olmadığı halde, her amino asit için ortalama olarak üç eş anlamlı kodon olduğu sonucuna varabiliriz.
Genlerin, DNA'nın çok uzun polinükleotid dizilerinin tekli uzantıları olarak tasarlandığı hatırlanacak olursa, tek genin başlangıcı ve bitişinin zorunlu olarak bellekte bulunması gerektiği açıktır.
PROTEİNLERİN BİYOSENTEZİ
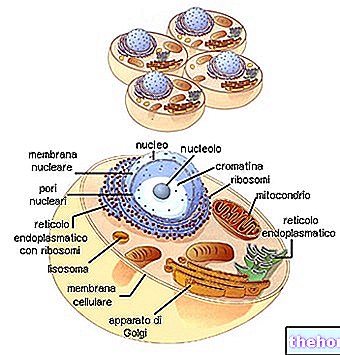
DNA'nın farklı bölümlerinde çift zincirin açılması ve farklı RNA türlerinin sentezi vardır.
Yükleme aşaması sırasında, RNAt amino asitlere bağlanır (önceden ATP ve spesifik enzim tarafından aktive edilir). Biyosentetik "makine", yanlış yüklenmiş tRNA'ları "düzeltemez".
RNAr daha sonra iki alt birime ayrılır ve ribozomal proteinlere bağlanarak ribozomların birleşmesini sağlar.
Sitoplazmadan geçen RNAm, ribozomlara bağlanır ve polisomu oluşturur.Mesajcı üzerinde akan her ribozom, amino asitleri alarak ve onları oluşum halinde polipeptit zincirine bağlayarak, ilgili kodonlara tamamlayıcı olan RNAt'ı yavaş yavaş barındırır.
Nispeten kararlı RNAt dolaşıma yeniden girer. Ribozomlar da tekrar kullanılır ve halihazırda monte edilmiş polipeptidi serbest bırakır.
Haberci, tümü monokatener olduğu için daha az kararlıdır, (ribonükleaz tarafından) kurucu ribonükleotidlere bölünür.
Böylece döngü, transkripsiyon tarafından sağlanan haberci RNA'lar üzerindeki polipeptitleri birbiri ardına sentezleyerek devam eder.